细胞如何在发育、稳态、损伤修复与疾病演进中决定命运轨迹?在复杂的组织微环境中,哪些细胞驱动了组织再生,又是哪些克隆在肿瘤复发或衰老过程中发生选择性扩增?这些疑问共同聚焦于生命科学的核心问题:如何构建细胞“过去”与“现在”的联系,进而预测其“未来”的命运,最终从细胞谱系演化的视角揭示细胞命运决定的底层规律。
5月29日,国际学术期刊Cell Stem Cell在线发表了国科大杭州高等研究院/中国科学院分子细胞科学卓越创新中心周斌团队与西湖大学王寿文团队合作撰写的特邀长篇综述,题为“Evolving Strategies for Lineage Tracing: Genetic Markers, Synthetic Barcodes, and Natural Variants”。该文聚焦现代谱系示踪领域的三大方法学支柱——遗传标记、合成条形码与自然变异,系统解析了各类技术的底层原理、适用场景、优势及局限。此外,文章深入探讨了谱系信息与单细胞多组学、空间图谱及计算模型的整合策略,为研究人员基于特定生物学问题设计谱系示踪实验提供了全景式指南(图1)。

图 1. 谱系示踪技术发展:遗传标记、合成条形码与自然变异
从“命运景观”到可量化框架
细胞命运图谱勾勒细胞从起源至终末身份的发育轨迹,深刻主导着组织稳态、再生修复与疾病发生。经典的Waddington表观遗传景观将细胞分化生动隐喻为沿起伏地形的滑落;而要将这一隐喻转化为可量化的科学框架,必须系统解析五大核心要素:细胞状态、命运路径、细胞功能输出、命运驱动力以及命运操控(图 2)。近年来,在单细胞测序、基因编辑、空间多组学与计算科学的交叉融合下,谱系示踪已突破传统观察性标记的局限,发展为集多维度、高通量与功能扰动于一体的前沿体系。该体系将上述五大要素深度整合,不仅揭示了“细胞当下处于何种状态”, 更精准重构了“细胞从何而来、历经何种命运分岔、因何锁定终末身份,以及最终发挥何种功能”。
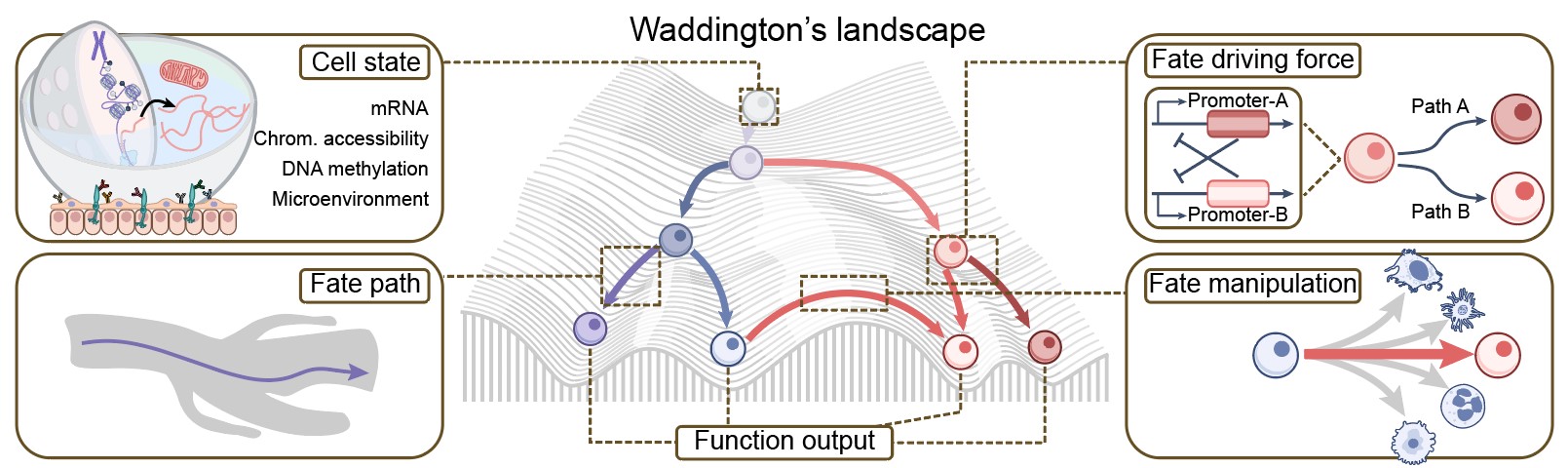
图 2. 细胞命运的可量化分析框架
遗传标记谱系示踪:从经典工具到高精度命运解析
基于遗传标记的谱系示踪是体内细胞命运研究的经典策略。该系统通常利用Cre、Flp、Dre等位点特异性重组酶激活荧光报告基因,从而对特定细胞群体进行永久标记,并在发育、稳态、损伤修复或疾病模型中追踪其后代。诱导型重组酶系统(如CreER)进一步赋予了该系统时间维度的可控性,使研究者能够将细胞标记精确锁定于特定发育窗口或病理阶段。近年来,针对单一标记难以精准靶向稀有、过渡态或高度动态细胞群体的局限,双重组酶与逻辑门系统显著提升了靶向特异性:AND逻辑门可精准捕获双阳性细胞,OR逻辑门有效降低背景泄漏,序贯重组策略则能更真实地模拟多步骤生物学过程并实现多层次功能解析,还能实时记录并捕获细胞的瞬时中间状态。此外,Brainbow、Confetti等随机多色报告系统以及MADM(双标记嵌合分析)技术,将研究分辨率从群体水平推进至单克隆水平,使量化单个前体细胞后代的异质性与功能差异成为可能(图 3)
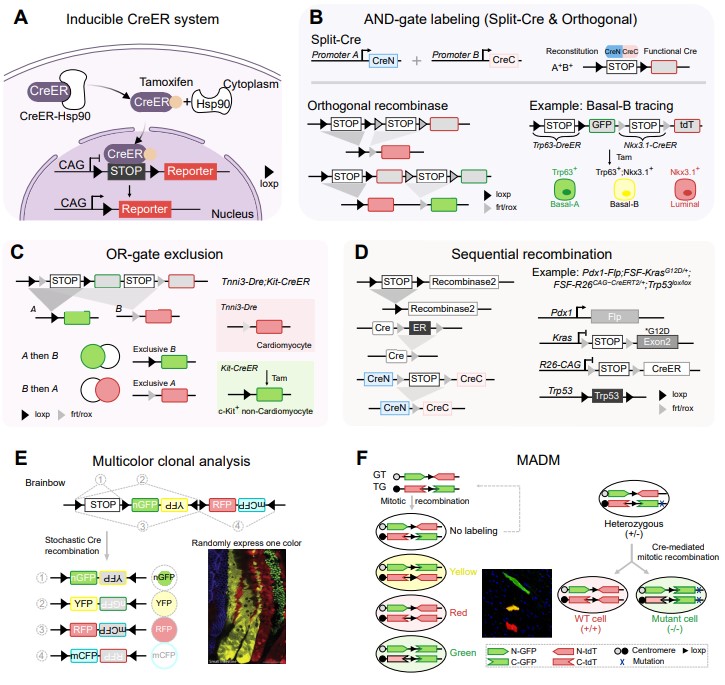
图 3. 基于遗传标记的谱系示踪与克隆分析策略
合成条形码谱系示踪:单细胞分辨率下大规模解析克隆命运
当研究旨在同时追踪海量克隆、重构复杂分化路径或捕捉早期命运偏向时,合成条形码技术提供了高效的高通量解决方案。静态条形码主要通过慢病毒文库等方式为细胞植入稳定DNA标签,结合单细胞测序可揭示命运偏向;整合可诱导报告模块的系统,还支持在活细胞层面分选目标克隆以进行功能探究。然而,病毒标记多局限于体外或移植模型,难以反映天然微环境下的细胞命运。尽管胚胎期宫内微注射部分缓解了此局限,广泛的原位克隆追踪仍主要依赖内源性静态条形码策略。其代表性系统包括转座酶(如 Sleeping Beauty、FlipJump)和重组酶介导的DNA重排(如 Polylox、LoxCode)。更重要的是,可进化条形码技术借助CRISPR/Cas9、末端脱氧核苷酸转移酶(TdT)辅助条形码插入、碱基或先导编辑等工具,在靶序列中持续累积可溯源的分子突变记录,从而使高分辨率重构连续的细胞谱系树成为可能。
需要强调的是,合成条形码技术的成功应用高度依赖于严谨的实验设计与算法校正:条形码homoplasy、单细胞测序信号的dropout、靶点缺失、编辑偏好性以及谱系树重构的不确定性,均会显著干扰克隆关系的推断。因此,必须将条形码复杂度、标记时机、采样规模、数据读出平台与建树算法作为一个整体进行系统性协同设计(图 4)。
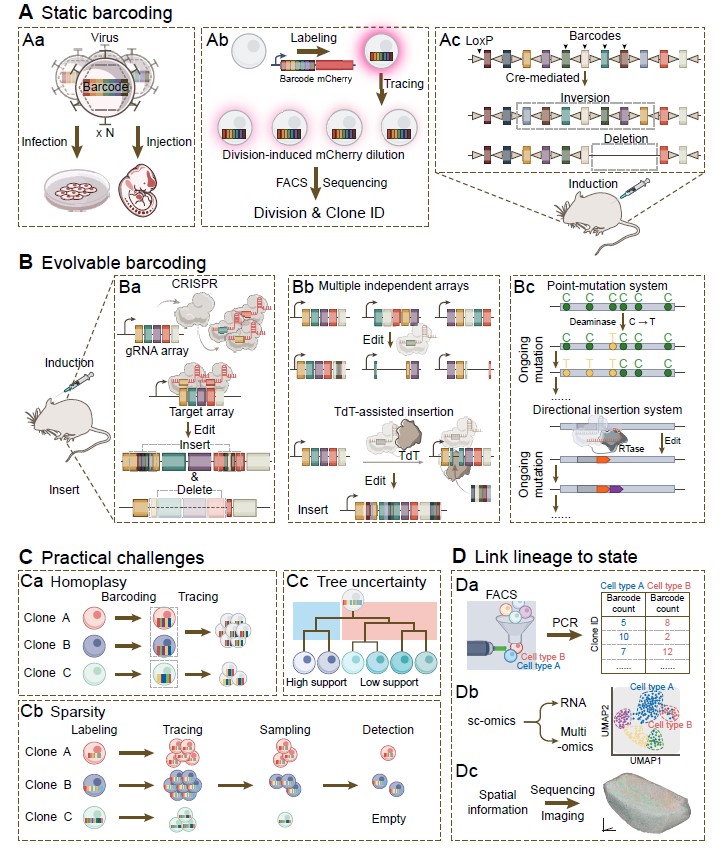
图 4. 合成条形码谱系示踪技术的应用和实践挑战
自然变异谱系示踪:无需预先标记的人体细胞历史回溯
在人体组织、临床样本以及其他无法进行转基因操作的体系中,利用细胞分裂过程中自然积累的内源性变异进行回溯性谱系重建,正在成为理解真实人类细胞命运的重要方法。核DNA体细胞单核苷酸变异(SNVs)具有较高保真度,结合全基因组测序或原代模板定向扩增(PTA)等技术,可用于重构人类胚胎发育、成体造血和疾病演进中的克隆结构。线粒体DNA变异因拷贝数高、突变率高,较易被常规单细胞平台捕获,适合解析部分短期或局部克隆事件。同时,DNA甲基化等稳定表观变异则为非侵入性谱系示踪提供了更丰富的信息库,MethylTree、EPI-Clone等方法推动了基于甲基化表观突变的人体细胞谱系树重建。与前两类策略相比,自然变异示踪的优势在于无需预先标记、适用于人类样本并保留天然细胞状态;其挑战则在于缺乏时间控制、突变率与测序深度之间存在成本与分辨率的取舍,并需要区分中性历史记录与具有功能意义的分子变化(图 5)。
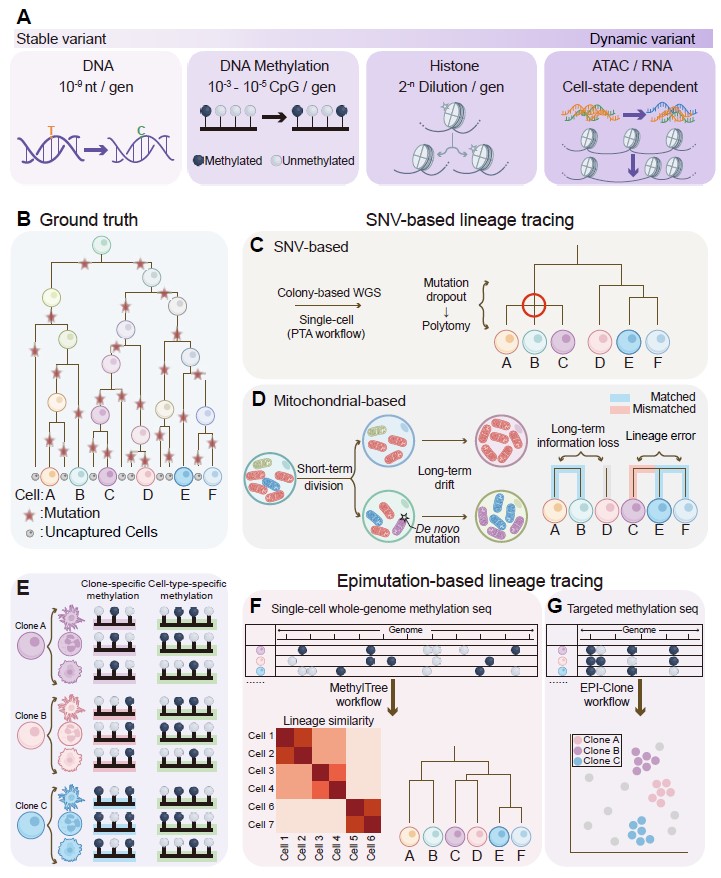
图 5. 基于自然发生的内源变异的谱系示踪策略
从谱系记录走向多维状态映射
单一谱系树能够揭示细胞之间的亲缘关系,却不足以完整理解命运决定。现代谱系示踪正在与单细胞多组学和空间组学深度融合:CellTag-multi、Camellia-seq 等平台可同步读取谱系条形码与转录组、染色质可及性或DNA甲基化信息;Visium、Stereo-seq 等空间转录组技术以及 intMEMOIR、PEtracer等原位读出策略,则使研究者能够在保留组织空间结构和微环境信息的同时解析克隆关系。面对高通量且常常稀疏的数据,计算方法同样成为关键支撑。CoSpar、LineageOT、moslin 等算法通过整合克隆共享、时间采样和转录组状态推断细胞转移概率;clone2vec、Clonotrace 等方法则从“克隆空间”组织大规模克隆行为。下一阶段的关键不只是记录更多谱系信息,而是建立能够同时整合谱系、状态、空间位置和功能扰动结果的统一分析框架。
面向实验设计与临床转化的指南
现代谱系示踪的三大支柱在标记特异性、克隆分辨率、时空控制和样本适用性方面各有取舍。遗传标记适合从明确起点出发,在体内追踪特定细胞群体并与基因敲除、细胞消融等功能扰动结合;合成条形码适合在可遗传操作的模型体系中开展大规模、单细胞分辨率的克隆命运解析;自然变异则为人类样本和临床研究提供了无需预先干预的回溯性方案。研究者在选择技术路线时,应首先明确核心问题:是要追踪某一特定细胞群体的命运,还是无偏解析大量克隆的分化潜能;是更重视空间结构保留,还是更重视系统发育树深度;是用于模型动物实验,还是面向人体样本和临床监测。
展望未来,谱系示踪将不仅服务于器官发育与组织再生机制研究,也有望进一步进入肿瘤演化、组织衰老、治疗耐受和克隆性造血等疾病场景。通过液体活检无创追踪体细胞突变和表观突变,未来或可实现癌症早筛、微小残留病灶监测、耐药克隆预警以及衰老人群克隆性造血风险分层。随着记录精度、空间分辨率和计算模型持续提升,谱系示踪有望从“回顾细胞历史”的工具,发展为预测和干预细胞命运的重要技术平台。
论文信息
国科大杭州高等研究院博士后康志鑫、中国科学院分子细胞科学卓越创新中心博士研究生陈惠与西湖大学科研助理李思阳为共同第一作者。国科大杭州高等研究院/中国科学院分子细胞科学卓越创新中心周斌研究员与西湖大学王寿文研究员为该综述的共同通讯作者。该研究获得国家重点研发计划、国家自然科学基金、中国科学院青年科学家基础研究项目、国科大杭州高等研究院科研基金以及新基石科学基金会等项目支持。
原文链接
https://www.cell.com/cell-stem-cell/fulltext/S1934-5909(26)00193-1
